Impairment of paravascular clearance pathways in the aging brain
- PMID: 25204284
- PMCID: PMC4245362
- DOI: 10.1002/ana.24271
Impairment of paravascular clearance pathways in the aging brain
Abstract
Objective: In the brain, protein waste removal is partly performed by paravascular pathways that facilitate convective exchange of water and soluble contents between cerebrospinal fluid (CSF) and interstitial fluid (ISF). Several lines of evidence suggest that bulk flow drainage via the glymphatic system is driven by cerebrovascular pulsation, and is dependent on astroglial water channels that line paravascular CSF pathways. The objective of this study was to evaluate whether the efficiency of CSF-ISF exchange and interstitial solute clearance is impaired in the aging brain.
Methods: CSF-ISF exchange was evaluated by in vivo and ex vivo fluorescence microscopy and interstitial solute clearance was evaluated by radiotracer clearance assays in young (2-3 months), middle-aged (10-12 months), and old (18-20 months) wild-type mice. The relationship between age-related changes in the expression of the astrocytic water channel aquaporin-4 (AQP4) and changes in glymphatic pathway function was evaluated by immunofluorescence.
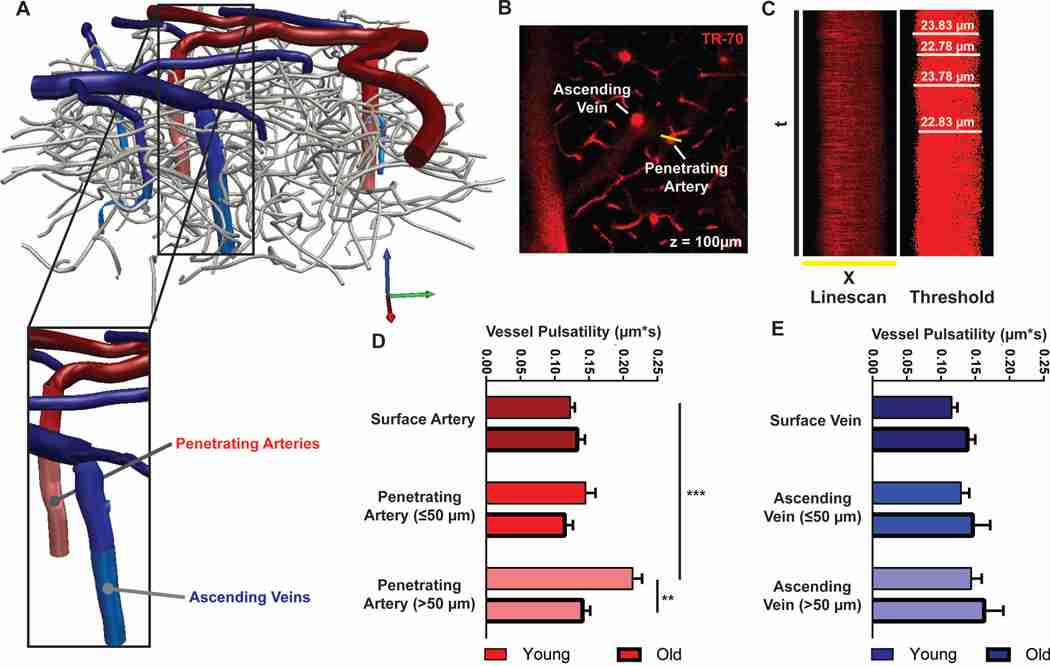
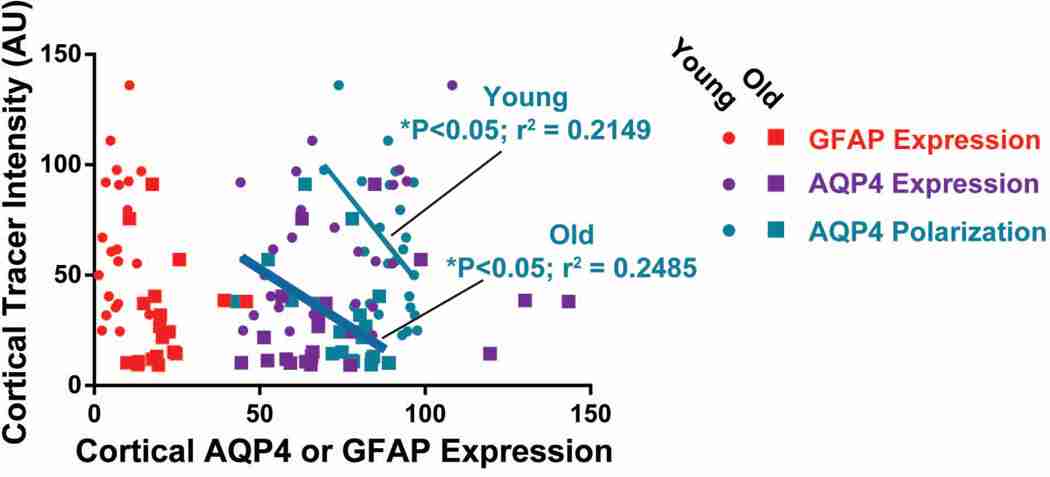
Results: Advancing age was associated with a dramatic decline in the efficiency of exchange between the subarachnoid CSF and the brain parenchyma. Relative to the young, clearance of intraparenchymally injected amyloid-β was impaired by 40% in the old mice. A 27% reduction in the vessel wall pulsatility of intracortical arterioles and widespread loss of perivascular AQP4 polarization along the penetrating arteries accompanied the decline in CSF-ISF exchange.
Interpretation: We propose that impaired glymphatic clearance contributes to cognitive decline among the elderly and may represent a novel therapeutic target for the treatment of neurodegenerative diseases associated with accumulation of misfolded protein aggregates.
© 2014 American Neurological Association.
Conflict of interest statement
Figures
Similar articles
-
Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain.J Neurosci. 2013 Nov 13;33(46):18190-9. doi: 10.1523/JNEUROSCI.1592-13.2013. J Neurosci. 2013. PMID: 24227727 Free PMC article.
-
A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β.Sci Transl Med. 2012 Aug 15;4(147):147ra111. doi: 10.1126/scitranslmed.3003748. Sci Transl Med. 2012. PMID: 22896675 Free PMC article.
-
The role of brain barriers in fluid movement in the CNS: is there a 'glymphatic' system?Acta Neuropathol. 2018 Mar;135(3):387-407. doi: 10.1007/s00401-018-1812-4. Epub 2018 Feb 10. Acta Neuropathol. 2018. PMID: 29428972 Review.
-
Voluntary Exercise Promotes Glymphatic Clearance of Amyloid Beta and Reduces the Activation of Astrocytes and Microglia in Aged Mice.Front Mol Neurosci. 2017 May 19;10:144. doi: 10.3389/fnmol.2017.00144. eCollection 2017. Front Mol Neurosci. 2017. PMID: 28579942 Free PMC article.
-
The Paravascular Pathway for Brain Waste Clearance: Current Understanding, Significance and Controversy.Front Neuroanat. 2017 Nov 7;11:101. doi: 10.3389/fnana.2017.00101. eCollection 2017. Front Neuroanat. 2017. PMID: 29163074 Free PMC article. Review.
Cited by 247 articles
-
Echocardiographic index E/e' in association with cerebral white matter hyperintensity progression.PLoS One. 2020 Jul 27;15(7):e0236473. doi: 10.1371/journal.pone.0236473. eCollection 2020. PLoS One. 2020. PMID: 32716979 Free PMC article.
-
The Lymphatic Vasculature in the 21st Century: Novel Functional Roles in Homeostasis and Disease.Cell. 2020 Jul 23;182(2):270-296. doi: 10.1016/j.cell.2020.06.039. Cell. 2020. PMID: 32707093 Review.
-
Retinal Vascular Pathology in a Rat Model of Cerebral Small Vessel Disease.Front Neurol. 2020 Jun 30;11:533. doi: 10.3389/fneur.2020.00533. eCollection 2020. Front Neurol. 2020. PMID: 32695061 Free PMC article.
-
Analyzing microglial-associated Aβ in Alzheimer's disease transgenic mice with a novel mid-domain Aβ-antibody.Sci Rep. 2020 Jun 29;10(1):10590. doi: 10.1038/s41598-020-67419-2. Sci Rep. 2020. PMID: 32601313 Free PMC article.
-
Altered brain fluid management in a rat model of arterial hypertension.Fluids Barriers CNS. 2020 Jun 26;17(1):41. doi: 10.1186/s12987-020-00203-6. Fluids Barriers CNS. 2020. PMID: 32590994 Free PMC article.
Publication types
MeSH terms
Substances
Grant support
LinkOut - more resources
-
Full Text Sources
-
Other Literature Sources
-
Medical

{kind=link}
{kind=link}