Molecular biology of the blood-brain and the blood-cerebrospinal fluid barriers: similarities and differences
- PMID: 21349151
- PMCID: PMC3045361
- DOI: 10.1186/2045-8118-8-3
Molecular biology of the blood-brain and the blood-cerebrospinal fluid barriers: similarities and differences
Abstract
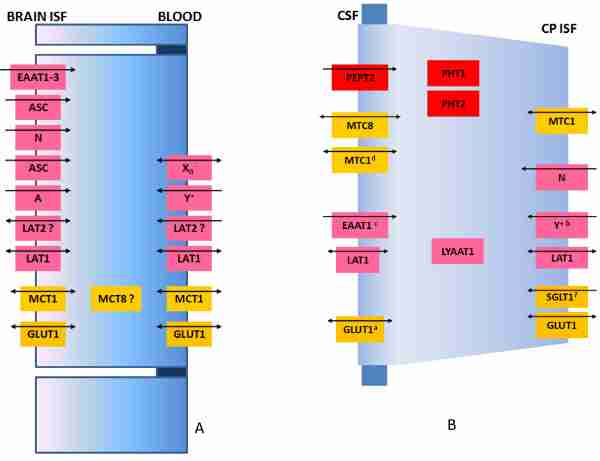
Efficient processing of information by the central nervous system (CNS) represents an important evolutionary advantage. Thus, homeostatic mechanisms have developed that provide appropriate circumstances for neuronal signaling, including a highly controlled and stable microenvironment. To provide such a milieu for neurons, extracellular fluids of the CNS are separated from the changeable environment of blood at three major interfaces: at the brain capillaries by the blood-brain barrier (BBB), which is localized at the level of the endothelial cells and separates brain interstitial fluid (ISF) from blood; at the epithelial layer of four choroid plexuses, the blood-cerebrospinal fluid (CSF) barrier (BCSFB), which separates CSF from the CP ISF, and at the arachnoid barrier. The two barriers that represent the largest interface between blood and brain extracellular fluids, the BBB and the BCSFB, prevent the free paracellular diffusion of polar molecules by complex morphological features, including tight junctions (TJs) that interconnect the endothelial and epithelial cells, respectively. The first part of this review focuses on the molecular biology of TJs and adherens junctions in the brain capillary endothelial cells and in the CP epithelial cells. However, normal function of the CNS depends on a constant supply of essential molecules, like glucose and amino acids from the blood, exchange of electrolytes between brain extracellular fluids and blood, as well as on efficient removal of metabolic waste products and excess neurotransmitters from the brain ISF. Therefore, a number of specific transport proteins are expressed in brain capillary endothelial cells and CP epithelial cells that provide transport of nutrients and ions into the CNS and removal of waste products and ions from the CSF. The second part of this review concentrates on the molecular biology of various solute carrier (SLC) transport proteins at those two barriers and underlines differences in their expression between the two barriers. Also, many blood-borne molecules and xenobiotics can diffuse into brain ISF and then into neuronal membranes due to their physicochemical properties. Entry of these compounds could be detrimental for neural transmission and signalling. Thus, BBB and BCSFB express transport proteins that actively restrict entry of lipophilic and amphipathic substances from blood and/or remove those molecules from the brain extracellular fluids. The third part of this review concentrates on the molecular biology of ATP-binding cassette (ABC)-transporters and those SLC transporters that are involved in efflux transport of xenobiotics, their expression at the BBB and BCSFB and differences in expression in the two major blood-brain interfaces. In addition, transport and diffusion of ions by the BBB and CP epithelium are involved in the formation of fluid, the ISF and CSF, respectively, so the last part of this review discusses molecular biology of ion transporters/exchangers and ion channels in the brain endothelial and CP epithelial cells.
Figures
Similar articles
-
The blood-brain and the blood-cerebrospinal fluid barriers: function and dysfunction.Semin Immunopathol. 2009 Nov;31(4):497-511. doi: 10.1007/s00281-009-0177-0. Epub 2009 Sep 25. Semin Immunopathol. 2009. PMID: 19779720 Review.
-
Fluid and ion transfer across the blood-brain and blood-cerebrospinal fluid barriers; a comparative account of mechanisms and roles.Fluids Barriers CNS. 2016 Oct 31;13(1):19. doi: 10.1186/s12987-016-0040-3. Fluids Barriers CNS. 2016. PMID: 27799072 Free PMC article. Review.
-
Cerebrospinal Fluid, Brain Electrolytes Balance, and the Unsuspected Intrinsic Property of Melanin to Dissociate the Water Molecule.CNS Neurol Disord Drug Targets. 2018;17(10):743-756. doi: 10.2174/1871527317666180904093430. CNS Neurol Disord Drug Targets. 2018. PMID: 30179148 Review.
-
SLC and ABC Transporters: Expression, Localization, and Species Differences at the Blood-Brain and the Blood-Cerebrospinal Fluid Barriers.AAPS J. 2017 Sep;19(5):1317-1331. doi: 10.1208/s12248-017-0110-8. Epub 2017 Jun 29. AAPS J. 2017. PMID: 28664465 Free PMC article. Review.
-
Physiology of blood-brain interfaces in relation to brain disposition of small compounds and macromolecules.Mol Pharm. 2013 May 6;10(5):1473-91. doi: 10.1021/mp300518e. Epub 2013 Jan 23. Mol Pharm. 2013. PMID: 23298398 Review.
Cited by 85 articles
-
Nanocarriers as Potential Drug Delivery Candidates for Overcoming the Blood-Brain Barrier: Challenges and Possibilities.ACS Omega. 2020 Jun 1;5(22):12583-12595. doi: 10.1021/acsomega.0c01592. eCollection 2020 Jun 9. ACS Omega. 2020. PMID: 32548442 Free PMC article. Review.
-
Personalized whole-body models integrate metabolism, physiology, and the gut microbiome.Mol Syst Biol. 2020 May;16(5):e8982. doi: 10.15252/msb.20198982. Mol Syst Biol. 2020. PMID: 32463598 Free PMC article.
-
Choroid plexus and the blood-cerebrospinal fluid barrier in disease.Fluids Barriers CNS. 2020 May 6;17(1):35. doi: 10.1186/s12987-020-00196-2. Fluids Barriers CNS. 2020. PMID: 32375819 Free PMC article. Review.
-
Neuroprotective effects of ammonium tetrathiomolybdate, a slow-release sulfide donor, in a rodent model of regional stroke.Intensive Care Med Exp. 2020 Apr 9;8(1):13. doi: 10.1186/s40635-020-00300-8. Intensive Care Med Exp. 2020. PMID: 32274608 Free PMC article.
-
Why do anti-inflammatory signals of bone marrow-derived stromal cells improve neurodegenerative conditions where anti-inflammatory drugs fail?J Neural Transm (Vienna). 2020 May;127(5):715-727. doi: 10.1007/s00702-020-02173-3. Epub 2020 Apr 6. J Neural Transm (Vienna). 2020. PMID: 32253536 Free PMC article. Review.
References
-
- Bradbury MW. The Concept of a Blood-Brain Barrier. Chichester: Wiley; 1979.
-
- Begley DJ, Brightman MW. Structural and functional aspects of the blood-brain barrier. Prog Drug Res. 2003;61:39–78. - PubMed
-
- Kuschinsky W, Paulson OB. Capillary circulation in the brain. Cerebrovasc Brain Metab Rev. 1992;4:261–286. - PubMed
LinkOut - more resources
-
Full Text Sources
-
Other Literature Sources
-
Miscellaneous

{kind=link}