Tamoxifen resistance and epigenetic modifications in breast cancer cell lines
- PMID: 18220739
- PMCID: PMC2789301
- DOI: 10.2174/092986707782794023
Tamoxifen resistance and epigenetic modifications in breast cancer cell lines
Abstract
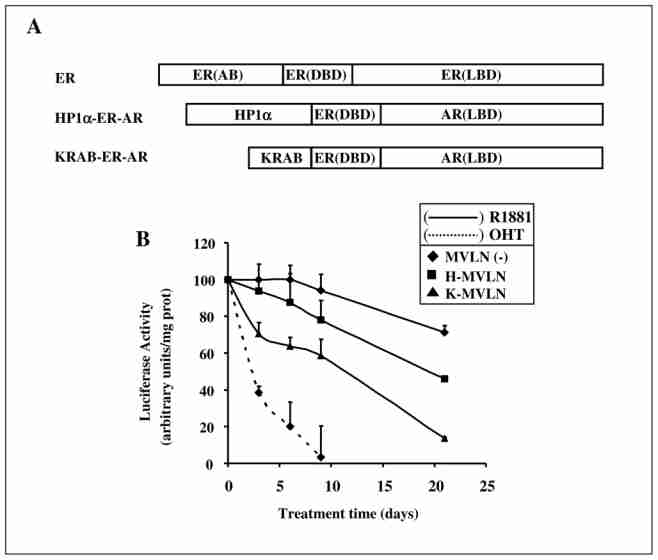
Epigenetic mechanisms play crucial roles in many processes, including neoplasia, genomic imprinting, gene silencing, differentiation, embryogenesis and X chromosome inactivation. Their relevance in human disease and therapy has grown rapidly with the recent emergence of drugs that target for example DNA methylation or histone acetylation. Epigenetic effects were also recently highlighted by the deciphering of the mechanism of action of steroid hormones and anti-hormones acting through nuclear receptors. In this review, we focus on the epigenetic effects associated with long-term treatment of breast cancer cells with the antiestrogen (AE) tamoxifen, in the context of resistance appearance. We summarize the data obtained with a model cell line developed in our laboratory supporting a role for HP1 proteins in the irreversible inactivation of gene expression by long-term treatment with AE.
Figures
Similar articles
-
Integrative analyses of gene expression and DNA methylation profiles in breast cancer cell line models of tamoxifen-resistance indicate a potential role of cells with stem-like properties.Breast Cancer Res. 2013 Dec 19;15(6):R119. doi: 10.1186/bcr3588. Breast Cancer Res. 2013. PMID: 24355041 Free PMC article.
-
Endocrine resistance in breast cancer: from cellular signaling pathways to epigenetic mechanisms.Transcription. 2012 Jul-Aug;3(4):165-70. doi: 10.4161/trns.20496. Epub 2012 Jul 1. Transcription. 2012. PMID: 22771991 Free PMC article. Review.
-
CtIP silencing as a novel mechanism of tamoxifen resistance in breast cancer.Mol Cancer Res. 2007 Dec;5(12):1285-95. doi: 10.1158/1541-7786.MCR-07-0126. Mol Cancer Res. 2007. PMID: 18171986
-
Epigenetic reactivation of estrogen receptor-α (ERα) by genistein enhances hormonal therapy sensitivity in ERα-negative breast cancer.Mol Cancer. 2013 Feb 4;12:9. doi: 10.1186/1476-4598-12-9. Mol Cancer. 2013. PMID: 23379261 Free PMC article.
-
Epigenetic regulation in estrogen receptor positive breast cancer--role in treatment response.J Mammary Gland Biol Neoplasia. 2010 Mar;15(1):35-47. doi: 10.1007/s10911-010-9166-0. Epub 2010 Jan 27. J Mammary Gland Biol Neoplasia. 2010. PMID: 20101445 Free PMC article. Review.
Cited by 18 articles
-
MiR-205 Dysregulations in Breast Cancer: The Complexity and Opportunities.Noncoding RNA. 2019 Nov 19;5(4):53. doi: 10.3390/ncrna5040053. Noncoding RNA. 2019. PMID: 31752366 Free PMC article. Review.
-
Dysregulation of DNA methylation patterns may identify patients with breast cancer resistant to endocrine therapy: A predictive classifier based on differentially methylated regions.Oncol Lett. 2019 Aug;18(2):1287-1303. doi: 10.3892/ol.2019.10405. Epub 2019 May 27. Oncol Lett. 2019. PMID: 31423189 Free PMC article.
-
The Central Contributions of Breast Cancer Stem Cells in Developing Resistance to Endocrine Therapy in Estrogen Receptor (ER)-Positive Breast Cancer.Cancers (Basel). 2019 Jul 22;11(7):1028. doi: 10.3390/cancers11071028. Cancers (Basel). 2019. PMID: 31336602 Free PMC article. Review.
-
Poly-ADP-Ribosylation of Estrogen Receptor-Alpha by PARP1 Mediates Antiestrogen Resistance in Human Breast Cancer Cells.Cancers (Basel). 2019 Jan 4;11(1):43. doi: 10.3390/cancers11010043. Cancers (Basel). 2019. PMID: 30621214 Free PMC article.
-
Bioinformatics-based identification of miR-542-5p as a predictive biomarker in breast cancer therapy.Hereditas. 2018 Jan 15;155:17. doi: 10.1186/s41065-018-0055-7. eCollection 2018. Hereditas. 2018. PMID: 29371858 Free PMC article.
Publication types
MeSH terms
Substances
LinkOut - more resources
-
Full Text Sources
-
Other Literature Sources
-
Medical
-
Miscellaneous

{kind=link}